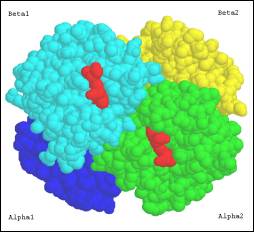
Alan Wardroper
Workshop: Introduction to Molecular Structure
Jan 13-16, 2004
Molecule:
Avian Oxy Haemoglobin (bar-headed goose, Anser indicus)
PDB Filename: 1a4f.pdb
X-ray Diffraction Resolution:
2.00 Å
Ref: Zhang
J, Hua Z, Tame JR, Lu G, Zhang R, Gu X. The crystal structure of a high oxygen affinity species of haemoglobin
(bar-headed goose haemoglobin in the oxy form). J. Mol. Biol. 1996 Jan 26; 255(3): 484-93.
General
Molecule: Avian Oxy Haemoglobin
Source: Anser indicus. Bar-headed goose
PDB Filename: 1a4f.pdb
(Zhang et al., 1996. The crystal structure of a high oxygen affinity species
of haemoglobin (bar-headed goose haemoglobin in the oxy form).
Introduction
The bar-headed goose Anser indicus has markedly higher tolerance for low oxygen pressures
than related lowland species, such as the greylag goose A.
anser, which is reflected in the significantly higher oxygen
affinity of its haemoglobin (Hb), allowing it to survive at higher altitudes. There
are only four amino acid differences between the Hbs of these two species. However, substitution
of Pro to Ala α119 in bar-headed goose Hb reduces contact between the α1
and β1 subunits as compared with A. anser, which confers increased oxygen affinity by destabilising
the deoxy T state of the protein as a result of the relaxation
of the α1β1 interface.
The crystal structure of A. indicus oxy-haemoglobin
was solved by X-ray diffraction at a resolution of 2 Å (PDB code, 1a4f;
Zhang et al., 1996) and is described here. The structure is also briefly
compared and contrasted with that of the deoxy T state (PDB code, 1hv4).
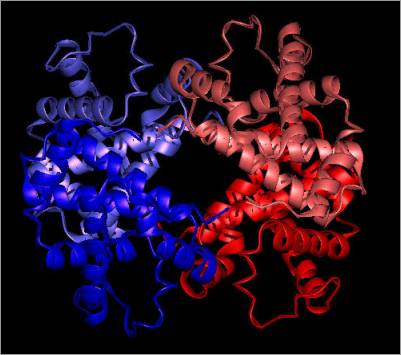
The bar-headed goose haemoglobin molecule
shows the same general overall structure as other Hbs characterised to date, consisting of a tetramer of two
α- and two β- chains of 141 and 146 residues, respectively, bound
to each other noncovalently. The structure is comprised primarily of antiparallel
helices (mostly α-helices) linked by short β-turns with no β-sheets
(Fig. 1).
Figure 1
Cartoon showing the tetrameric structure
of Avian Haemoglobin
The molecule is characterised according to the SCOP
classification as having a 6-helix, folded leaf, partly opened core belonging
to the globin-like
superfamily. Overall, the structure consists of 68.25% α-helix
(i→i+4) and 6.775% 310-helix (i→i+3)
by sequence across all chains, as listed in Table
1. There are also three short right-handed 310 helices,
characterised by tighter coiling with an i->i+3 Hydrogen bonding topology. The helices are connected
by loops and turns, and the molecule has no beta strands or disulfide bonds.
The unstructured loop regions lie mostly on the outside of the molecule, exposed
to the surrounding solvent, while the solvent-exposed helices are amphipathic,
with hydrophilic polar side chains exposed on the external face – (oxygen
and nitrogen are present on the external surface at high frequency – and hydrophobic
non-polar side chains facing into the hydrophobic core (Fig.
2 A & B). Figure 7 shows
Ramachandran plots indicating the mainly helical secondary structure in both
α- and β-chains. Residues βAla119 and αLeu2 have unusual
ψ and φ bond torsion angles as indicated in the plots.
Figure 2
Surface α-helices are amphipathic
| A)
External surface of α-helix |
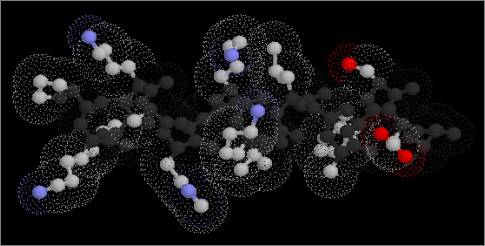
A) Side view of external face of α-helix
showing polar side chains exposed to the solvent. |
| B)
End view showing inner (left) and outer (right) faces |
| 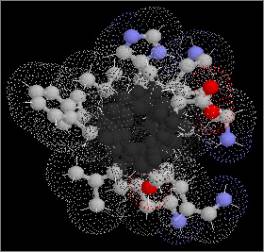
B) The other, internal face presents
non-polar side chains into the hydrophobic core of the molecule, as
shown in this end view: left, internal (hydrophobic); right, external
(hydrophilic). |
The structure of the avian Hb is similar to that of human HbA (1hhO;
Shaanan, 1983),
but differences in intersubunit contacts due to the single Pro→Ala mutation described above result in slightly altered
packing of the tetramer. In oxy Hb of Anser indicus, the αβ
dimers are rotated by an angle of about
4° as compared with human oxy Hb. This unique quaternary structure, designated the RH
state, may be correlated with the increased oxygen affinity of bar-headed
goose Hb described above. In the native folded state, the N-
and C-termini (Val/Arg and Val/His for α- and β-chains, respectively)
of each chain are localized in close proximity to each other and form a strongly
positively charged groove at the entrance to the central cavity. The distribution
of positive charges around this highly conformationally ordered region suggests
that this is the binding site for inositol pentaphosphate in the T state,
which binds avian Hb
and is required for function in the same way as 2,3-diphosphoglycerate binds
and regulates oxygen affinity of mammalian Hb (Rollerma & Bauer,
1979; Arnone, 1972).
Topology
The α-chain
(141 residues), consisting of seven α-helices and one 310
helix linked in antiparallel orientation mostly by β-turns, has the following
topology: HTHGTTHTHHTHTHTT (H=helix, T=turn, G=γ-turn). The 146-residue
β-chain consisting of seven α- and two 310-helices, has the following
topology: HTHGHHTHHGHT (Tables 1
& 2). Helix 3 in each chain is a right-handed
310 helix (α- and β-chains, residues 37-42 and 36-42,
respectively). The β-chain also has an additional short 310-helix
from residues 43-45 (Fig. 3, Tables 1 &
2).
Figure 3
PDB secondary structure cartoon
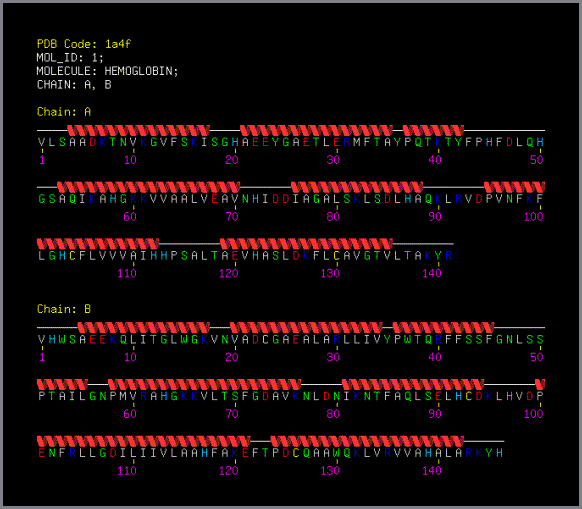
PDB secondary structure cartoon showing the
7 helices in the A chain and 8 in the B chain.
Generated by STING
Millennium Suite
Active sites
There are four active sites in the tetrameric
molecule, one per chain, each of which binds a haem group containing one Fe++
atom (which is oxidised in 1a4f) (Fig. 4).
The key residues for haem group binding are outlined in Table
3. In both α- and β-chains, the haem group is coordinated
between two basic His residues and a Lys residue. The proximal His residues, His87
and His92 in the α- and β-chain, respectively, make contact with
the haem group Fe++ ion, forming a ‘picket fence’ to stabilise
the ferrous form of the iron porphyrin ring necessary for reversible binding
of O2 for extended periods (Collman et al., 1975).
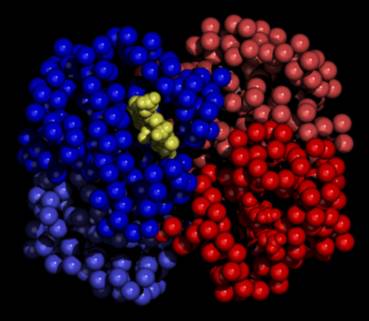
Figure 4
Haem binding clefts in the haemoglobin
tetramer
| Oxy Haemoglobin (Haem group shown) |
Oxy Haemoglobin (Haem group not shown) |
| 
|
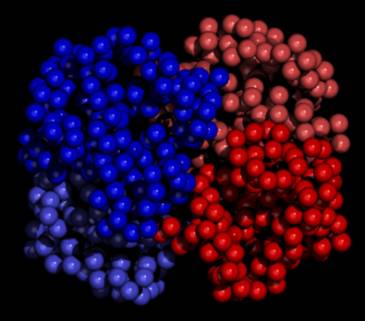
|
Space-filling models showing the haemoglobin
tetramer with (left) and without (right) haem groups bound.
The β1-chain is shown in
blue on the left and the α2-chain is shown in red on the
right.
Note, the models, generated using QUANTA,
are schematic in that they do not represent the physiological structure
in the absence of haem binding.
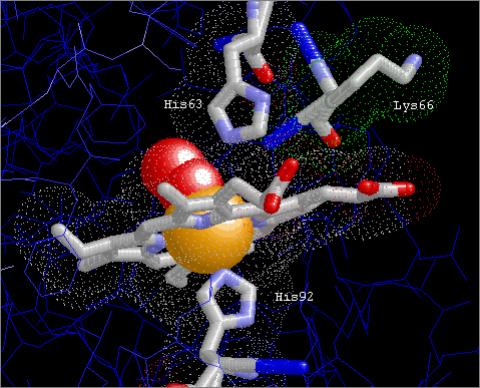
In the β-chain, Lys66 forms
a Hydrogen bond with the haem group via βLys66Nζ→Haem
Ala65 CβH over a distance of 3.294Å. The equivalent residue in the α-chain, Lys61, may also interact with
the haem group, although it is located at the greater distance of 3.57Å and
no H-bond was detected in the model.
Figures 5A
and B show the relative positions of the distal (αHis58; βHis63) and proximal
(βHis92; αHis87) His residues, βLys66, and the haem group in Hb the oxy R state. Figure 5B shows the
Van der Waal’s surfaces of the haem group and each of the three key residues.
βLys66Nζ can be seen to form an H-bond with the haem group Ala65
CβH (Figs. 5A & B).
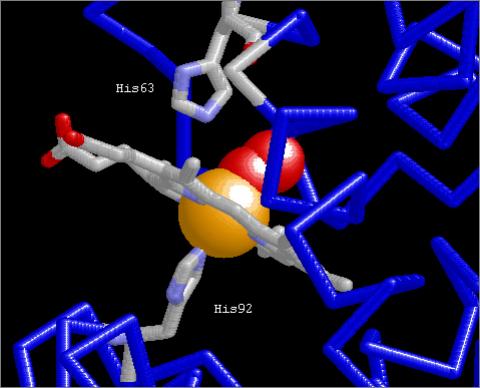
Figure 5
Haem binding cleft (β-chain) showing
contact with His92
| A)
His92 makes contact with Haem Fe |
B)
Van der Waal’s radii of contact area |
|
 |
 |
The proximal His residue (βHis92, αHis87)
stabilises the Fe++ ion in the iron porphyrin ring of the haem
group in R state haemoglobin. Figure 5A shows the locations
of the basic proximal (βHis92) and distal (βHis63) His residues
relative to the haem group. Figure 5B shows the Van
der Waal’s surfaces of the haeme group and the three key residues. βLys66Nζ
forms an H-bond with the haem group Ala65 CβH.
(Images generated using RasMol)
The hydrophilic propanoate groups
of the haem group face the water at the surface of the protein, while the
hydrophobic portions of the ring are buried among the hydrophobic amino acids
of the protein chains. Figure 6 shows
H2O molecules coordinated with the haem groups bound to the α-
and β-chains. As can be seen in the figure, the α-chain haem has two coordinated
water molecules, while that in the B chain has only 1, and they are located
in different positions relative to the porphyrin ring structure (which in
itself is altered between the two states, as described in the next section).
In addition, the haem groups can be seen to lie in different planes across
the chains, with a difference in angle of ~36° between α1
and β1 (Fig. 6).
Figure 6
Water molecules coordinated with haem
groups
The hydrophilic propanoate groups of the
haem group face the solvent at the surface of the protein, while the hydrophobic
portions of the ring are buried among the hydrophobic amino acids of the protein
chains. The α-chain haem carries two coordinated water molecules, while
that of the β-chain has only 1, and they are located in different positions
relative to the porphyrin ring structure.
Fig. 7
Ramachandran Plots
| α-Chain |
β-Chain |
|
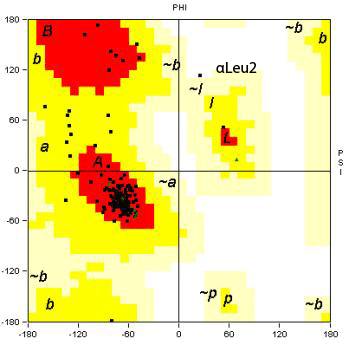 |
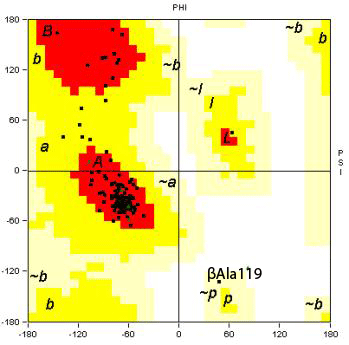 |
Plotted using STING
Ramachandran plots indicating the mainly
helical secondary structure in both α- and β-chains.
βAla119 and αLeu2 have unusual
torsion angles:
βAla119: PHI, 47.87;
PSI, -132.55
αLeu2: PHI, 25.75;
PSI, 113.78
Comparison between T (tense) and R (relaxed) states
Comparison of the oxy-Hb structure (1a4h) with the deoxy
structure (1hv4) for the same species
revealed differences in both the overall structure and the local conformation
of the haem group between the two states. In the oxy R form, the Fe atom is
lifted up and through the haem ring structure orthologous to the porphyrin
plane, which translates throughout the ring causing its shape to change. The
change in oxidation state of the haem group also translates to global shifts
in relative orientation of domains throughout the molecule. While the distance
between βHis63 and the ferrous iron atom changes relatively little with oxidation state [βHis
63↔Fe: Oxy (1a4f), 4.508Å; Deoxy (1hv4), 4.23Å], βHis92 is brought
into closer proximity with the haem Fe on O2 binding [βHis92Oxy↔Fe:
Oxy (1a4f), 3.046 Å, Deoxy (1hv4), 2.32Å]. As the Fe atom and βHis63
remain closely associated, this movement of the Fe atom on oxidation pulls
the His residue out of its T state position, which is translated to a shift
in the entire protein chain into the R state. These changes are transmitted
throughout the protein, ultimately resulting in the marked shift in shape
that alters the binding strength of the neighbouring sites. Clear differences
in overall structure can be seen in Figure 8,
which shows only the β-chains of both oxy and deoxy
forms.
Figure 8
Stereo
views showing differences in relative positions of haem Fe, βHis63, and
βHis92 between T and R states
|
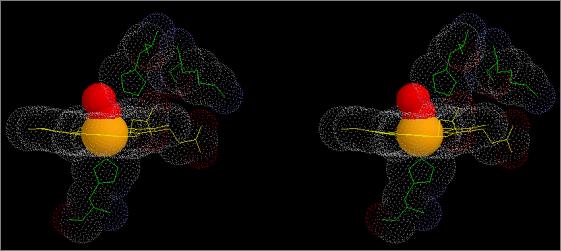 |
Oxy
(R) Haemoglobin (1a4f) |
|
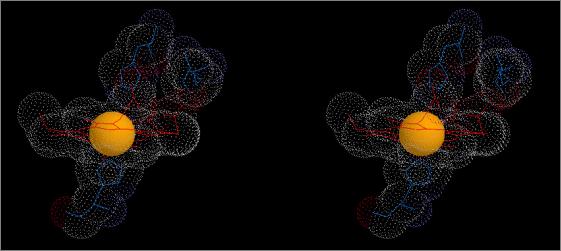 |
Deoxy
(T) Haemoglobin (1hv4) |
The figures show stereo views of the haem
binding pocket of A. indicus haemoglobin
in both R (oxy; top) and T (deoxy; bottom) states.
Note the differences in
relative positions of the Fe atom and the haem group. Note also the changes
in position and orientation of the proximal and distal His residues as well
the shift in position of the auxiliary Lys66 residue between T and R states.
As can be seen from Tables
4 and 5, the inter-atom
bond lengths deviate much less from the overall means in Hb in the deoxy
state as compared with the oxy state.
Although the difference in resolution of the structures will affect these
values (1a4f, 2.00 Å; 1hv4, 2.80 Å), these differences reflect the conformational
changes known to occur in Hb on changing between oxidation states (Baldwin & Chothia, 1979). The conformational changes that occur on binding
of oxygen to the haem Fe are communicated to the interfaces
between the subunits, breaking the salt links between the chains leading to
a change from the T to the R state on oxidation. Thus, in the deoxy- T, or ‘tense’, conformation the quaternary structure
is markedly more constrained than in the R, or ‘relaxed’, conformation. This
is especially noticeable at the C-termini of each chain, which have almost
full rotational freedom in the R state but are highly constrained by electrostatic
interactions in the T state.
Conclusions
The haemoglobin of the bar-headed goose Anser indicus is highly similar to mammalian
Hbs as well as those of other avian species both structurally and in its amino
acid sequence. The unusually high oxygen affinity of the Hb of this
species represents an adaptation to high altitudes, and appears to be due
to a single amino acid substitution of the bulky cyclic imino
acid residue Pro to the small aliphatic
residue Ala at position 119 in the α-chain, removing a hydrophobic contact
between the α1 and β1 subunits, resulting
in a loosening of the contact between the subunits as compared with the Hb from the related lowland species A. anser. This confers increased oxygen
affinity by destabilising the deoxy
T state of the protein as a result
of the reduced steric hindrance at the α1β1 interface.
References
- Arnone A. X-ray diffraction study of binding
of 2,3-diphosphoglycerate to human deoxyhaemoglobin. Nature. 1972 May
19; 237(5351): 146-9.
- Baldwin J, Chothia C. Haemoglobin: the structural
changes related to ligand binding and its allosteric mechanism. J Mol
Biol. 1979 Apr 5; 129(2): 175-220.
- Collman JP, Gagne RR, Reed CA, Halbert TR, Lang
G, Robinson WT. "Picket fence porphyrins." Synthetic models
for oxygen binding hemoproteins. J Am Chem Soc. 1975 Mar 19;
97(6): 1427-39.
- Liang, Y, Hua, Z, Liang, X, Xu, Q, Lu, G. The
Crystal Structure of Bar-Headed Goose Hemoglobin in Deoxy Form: The Allosteric
Mechanism of a Hemoglobin Species with High Oxygen Affinity. J.Mol.Biol.
313 pp. 123 (2001)
- Rollema HS, Bauer C. The interaction of inositol
pentaphosphate with the hemoglobins of highland and lowland geese. J
Biol Chem. 1979 Dec 10; 254(23): 12038-43.
- Shaanan B. Structure of human oxyhaemoglobin
at 2.1 Å resolution. J Mol Biol. 1983 Nov 25; 171(1): 31-59.
- Zhang J, Hua Z, Tame JR, Lu G, Zhang R, Gu X.
The crystal structure of a high oxygen affinity species of haemoglobin
(bar-headed goose haemoglobin in the oxy form). J. Mol. Biol. 1996 Jan
26; 255(3): 484-93.
Tables
Table 1::Comparison
of structural features between Oxy/Deoxy Hb
Table 2::ProMotif
Summary
Table 3::Key Residues
for Haem Binding
Table 4::Oxy Hb
Chain Bond Lengths
Table 5::Deoxy Hb
Chain Bond Lengths
Table 6::Oxy Haemoglobin
Helix Details
Table 7A::List
of putative hydrogen bonds formed by LYS 61 (α-chain) in PDB entry 1A4F
Table
7B::List
of putative hydrogen bonds formed by LYS 66 (β-chain) in PDB entry 1A4F
Table 7C::List
of putative hydrogen bonds formed by HIS 58 (a-chain) in PDB entry 1A4F
Table 7D::List
of putative hydrogen bonds formed by HIS 87 (α-chain) in PDB entry 1A4F
Table 8A::List
of putative hydrogen bonds formed by HIS 63 (β-chain) in PDB entry 1A4F
Table 8B::List
of putative hydrogen bonds formed by HIS 92 (β-chain) in PDB entry 1A4F
Table 9::PROCHECK
summary for 1a4f